
Скелет лягушка
Скелет земноводного
ТЕМА 10. СКЕЛЕТ ЗЕМНОВОДНЫХ СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ ОБЪЕКТА
Подтип Позвоночные, Vertebrata Класс Земноводные, Amphibia Отряд Бесхвостые, Anura (Ecaudata) Представитель — Озерная лягушка, Rana ridibunda Pall.
МАТЕРИАЛ И ОБОРУДОВАНИЕ
На одного-двух студентов необходимы: 1. Смонтированный на картонных планшетах разобранный скелет лягушки. 2. Препарировальные иглы — 2.
ЗАДАНИЕ
Уяснить особенности строения скелета земноводных. Сделать следующие рисунки: 1. Череп лягушки сверху. 2. Череп снизу. 3. Позвоночный столб и прикрепленный к нему тазовый пояс сверху. 4.Пояс передних конечностей (расправленный) снизу. 5. Скелет передней конечности. 6. Тазовый пояс сбоку. 7. Скелет задней конечности.
Дополнительное задание
Сравнить, не зарисовывая, скелеты бесхвостого (лягушка) и хвостатого земноводного (влажный препарат).
ОПИСАНИЕ СКЕЛЕТА
Скелет земноводных, как и других позвоночных животных, разделяется на осевой скелет (позвоночный столб), череп (мозговой и висцеральный), парные конечности и их пояса.
Практически во всех отделах скелета еще довольно большую роль играет хрящ. Осевой скелет. Осевой скелет у земноводных представлен позвоночным столбом (columna veriebralis; рис. 54),состоящим из окостеневших позвонков; хорда во взрослом состоянии обычно редуцируется. По сравнению с рыбами осевой скелет земноводных состоит из большего числа отделов.
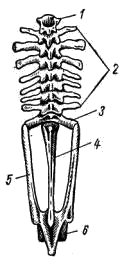
Рис. 54. Осевой скелет и тазовый пояс лягушки (вид сверху): 1 — шейный позвонок, 2 — туловищные позвонки, 3 — крестцовый позвонок. 4 — уростиль (слившиеся хвостовые позвонки), 5 — тазовый пояс, 6 — вертлужная впадина
1. Шейный отдел (pars cervicalis; рис. 54, 1) у всех земноводных представлен одним шейным позвонком, который при помощи двух суставных площадок подвижно сочленяется с черепом.
2. Туловищный отдел (pars thoracalis; рис. 54, 2) позвоночника лягушек состоит из 7 позвонков (у хвостатых земноводных — от 14 до 63).
3. Крестцовый отдел (pars sacralis; рис. 54, 3) у всех земноводных представлен одним крестцовым позвонком, к массивным поперечным отросткам которого причленяются подвздошные кости тазового пояса (рис. 54, 5).
4. Хвостовой отдел (pars caudalis) у личинок бесхвостых земноводных состоит из довольно большого числа отдельных позвонков, которые во время метаморфоза сливаются в одну хвостовую косточку — уростиль (urostyl; рис. 54, 4). У хвостатых земноводных в хвосте сохраняется 26—36 отдельных позвонков.
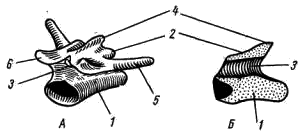
Рис. 55. Туловищный позвонок лягушки А — общий вид; Б — продольный разрез: 1 — тело позвонка, 2 — верхняя дуга, 3 — канал для спинного мозга, 4 — остистый отросток, 5 — поперечный отросток, 6 — сочленовный отросток
Туловищные позвонки большинства лягушек процельного типа: тело позвонка спереди вогнуто, сзади выпукло (рис. 55), однако последний туловищный позвонок имеет амфицельный (двояковогнутый) тип строения. Над телами позвонков располагаются верхние дуги (arcus neuralis; рис. 55, 2), образующие канал для спинного мозга. На спинной, стороне дуги имеется небольшой остистый отросток (processus spinosus; рис. 55, 4). От верхнебоковой поверхности тела позвонка отходят парные поперечные отростки (processus transversus; рис. 55, 5); у хвостатых земноводных к их концам прикрепляются коротенькие ребра, у бесхвостых земноводных ребра отсутствуют. Позвонки соединяются друг с другом сочленением самих тел позвонков (что обеспечивается процельным типом их строения) и соединением специальных парных сочленовных отростков (processus atricularis; рис. 55, 6), располагающихся спереди и сзади на основании верхней дуги.
По сравнению с рыбами для земноводных характерна большая дифференцировка позвоночного столба на отделы, изменение формы тел позвонков и более сильное развитие сочленовных отростков. Эти преобразования связаны с наземным образом жизни и обеспечивают большую прочность осевого скелета при сохранении его подвижности, прочное соединение с ним тазового пояса и допускают некоторую подвижность черепа в вертикальной плоскости относительно туловища (возможность поднимать и опускать голову). Череп. Осевой, или мозговой, череп земноводных, как и череп хрящевых рыб, платибазального типа: с широким основанием и широко расставленными глазницами, между которыми располагается передний конец головного мозга. В черепе, по сравнению с костистыми рыбами, сохраняется много хряща, а число окостенений относительно невелико.
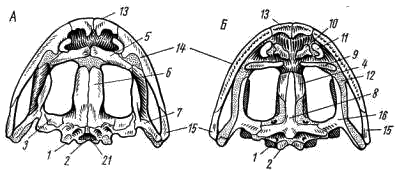
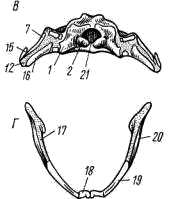
Рис. 56. Череп лягушки А — сверху; Б — снизу; В — сзади; Г — нижняя челюсть сверху, пунктиром показаны хрящевые участки черепа 1 — боковая затылочная кость, 2 — затылочный мыщелок, 3 — переднеушная кость, 4 — клиновидно-обонятельная кость, 5 — носовая кость, 6 — лобно-теменная кость, 7 — чешуйчатая кость, 8 — парасфеноид, 9 — нёбная кость, 10 — сошник, 11 — хоана, 12 — нёбно-квадратный хрящ, 13 — межчелюстная кость, 14 — верхнечелюстная кость, 15 — квадратно-скуловая кость, 16 — крыловидная кость, 17 — меккелев хрящ, 18 — подбородочно-челюстная кость, 19 — зубная кость, 20 — угловая кость, 21 — большое затылочное отверстие
В хряще затылочного отдела мозгового черепа образуются только парные боковые затылочные кости (occipitale laterale; рис. 56, 1), окаймляющие большое затылочное отверстие (foramen occipitale magnum; рис. 56, 21). Каждая из них образует мыщелок (condylus occipitalis; рис. 56, 2) для сочленения с шейным позвонком. В области слуховой капсулы вместо пяти пар ушных костей, характерных для костистых рыб, у земноводных возникает лишь одна пара — переднеушные кости (prooticum; рис. 56, 3). В передней части мозгового черепа при окостенении хряща образуется непарная клиновиднообонятельная кость (sphenethmoideum, рис. 56, 4), имеющая вид костного кольцевого пояска.

Рис. 57. Хрящевой череп головастика: 1 — мозговой череп, 2 — нёбно-квадратный хрящ, 3 — меккелев хрящ, 4 — жаберные дуги, 5 — челюстной сустав
Вся остальная часть мозгового черепа остается хрящевой. Ее укрепляют покровные (кожные) кости. Сверху в передней части черепа лежат имеющие удлиненно- треугольную форму парные носовые кости (nasale; рис. 56, 5), затем слившиеся из лобных и теменных костей парные лобно-теменные кости (frontoparietale, рис. 56, 6), и кнаружи от ушных костей — имеющие сложную форму чешуйчатые кости (squamosum; рис. 56, 7). Дно мозгового черепа прикрывает мощная покровная кость крестообразной формы — парасфеноид (parasphenoideum; рис. 56, 8). Спереди от него лежат тоже покровные парные нёбные кости (palatinum; рис. 56, 9) и парные сошники (vomer; рис. 56, 10); на сошниках сидят мелкие зубы. Впереди сошников располагаются парные внутренние ноздри — хоаны (рис. 56, 11).
Висцеральный отдел черепа земноводных также сохраняет много хряща. В течение всей жизни сохраняется нёбно-квадратный хрящ (cartilago palatoquadratum; рис. 56, 12), прирастающий передним концом к обонятельной области мозгового черепа, а задним — к основанию черепа перед слуховой капсулой (рис. 57, 2). Поэтому череп земноводных, как и остальных наземных позвоночных животных, по типу прикрепления челюстной дуги — аутостилический.
К нёбно-квадратному хрящу прилегают возникающие в коже кости вторичной верхней челюсти: парные межчелюстные кости (intermaxillare или praemaxillare; рис. 56, 13), несущие зубы и верхнечелюстные кости (maxillare; рис. 56, 14). За ними, укрепляя заднюю часть нёбно-квадратного хряща, сверху образуется покровная квадратно- скуловая кость (quadratojugale; рис. 56, 15), а снизу также покровная — крыловидная кость (pterygoideum; рис. 56, 16).
Первичная нижняя челюсть — меккелев хрящ (cartilago Meckeli; рис. 56, 17) также остается хрящевой, только самый ее передний конец окостеневает в маленькие парные подбородочно-челюстные кости (mento-mandibulare, рис.56, 18). К ним присоединяются покровные зубные кости (dentale; рис. 56, 19), у земноводных лишенные зубов. Задняя часть меккелева хряща обрастает длинной покровной угловой костью (angulare; рис. 56, 20) и еще несколькими мелкими покровными косточками. Через суставной отросток меккелева хряща нижняя челюсть подвижно сочленяется с задней частью нёбно- квадратного хряща (рис. 57, 5).
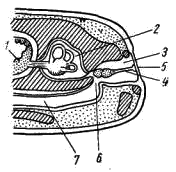
Рис. 58. Схематический разрез через слуховую область головы лягушки: 1 — головной мозг, 2 —слуховая капсула с полукружными каналами, 3 — полость среднего уха, 4 — стремечко, 5 — барабанная перепонка, 6 — евстахиева труба, 7 — ротовая полость
Полная редукция у земноводных жаберной крышки и смена гиостилического типа прикрепления челюстей аутостилистическим приводят к потере основных функций подъязычной дуги (укрепление челюстей, опора жаберной крышки).
Подъязычная дуга еще у предков современных земноводных начала редуцироваться, а полость брызгальца (остаток жаберной щели между челюстной и подъязычной дугами) в связи с переходом к жизни в воздушной среде преобразовалась в полость среднего уха (рис. 58, 3). Расположенный рядом с брызгальцем верхний элемент подъязычной дуги — подвесок (hyomandibulare) превратился в слуховую косточку—столбик, или стремечко (columel1а, или stapes; рис. 58, 4). У современных бесхвостых земноводных стремечко имеет вид тоненькой палочковидной косточки, лежащей перпендикулярно мозговому черепу под чешуйчатой и квадратно-скуловой костями. Одним концом стремечко упирается в центр барабанной перепонки (рис. 58, 5), а другим — в овальное окно слуховой капсулы. Этот механизм, усиливающий звуковые колебания и обеспечивающий возможность слуха в воздушной среде, у части современных земноводных в различной степени вторично редуцирован. Добавочными механизмами, обеспечивающими восприятие звуковых волн, распространяющихся по твердому субстрату, у них становится нижняя челюсть, а также передача звуковых колебаний по кровеносным стволам.

Рис. 59. Подъязычный аппарат лягушки: 1 — тело, 2 — рожки
Нижний элемент подъязычной дуги — гиоид (hyoideum) и функционирующие у личинок земноводных жаберные дуги во время метаморфоза превращаются в подъязычный аппарат (рис. 59). У бесхвостых земноводных он представляет собой хрящевую пластинку с двумя главными нарами отростков — рожек. Передние, более длинные рожки (видоизменившиеся гиоиды) направляются назад и вверх и прикрепляются к стенкам слуховых капсул мозгового черепа. Подъязычный аппарат укрепляет дно ротовой полоски: к нему прикрепляются мышцы, расположенные между ветвями нижней челюсти.
Предполагают, что гортанные хрящи также представляют собой преобразованные остатки жаберных дуг. Парные конечности и их пояса. Конечности земноводных, кяк и конечности других классов наземных позвоночных животных, представляют в схеме систему рычагов, подвижно соединенных друг с другом. Схемы строения передней и задней конечностей однотипны (рис. 60):
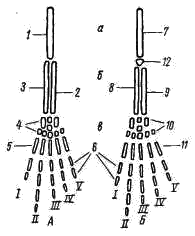
Рис. 60. Схема строения парных конечностей (левых) наземных позвоночных А — передняя конечность; Б — задняя конечность: а — плечо — бедро, б — предплечье — голень, а — кисть—стопа; 1 — плечевая кость, 2 — локтевая кость, 3 — лучевая кость, 4 — запястье, 5 — пясть, 6 — фаланги пальцев, 7 — бедренная кость, 8 — большая берцовая кость, 9 — малая берцовая кость, 10 — предплюсна, 11 — плюсна, 12 — коленная чашечка, I — V — пальцы
Проксимальный конец имеет округлую головку плеча (caput humeri; рис. 61, 2), которая входит в суставную ямку пояса передних конечностей; на дистальном конце — полушаровидная поверхность для сочленения с костями предплечья. Поверхность плеча имеет гребни, к которым прикрепляются мышцы. У бесхвостых земноводных лежащая снаружи локтевая (ulna; рис. 61, 4) и с внутренней стороны — лучевая (radius; рис. 61, 5) кости сливаются в единую кость предплечья (antebrachium, рис. 61, 3); продольная бороздка показывает границу их слияния. У хвостатых земноводных эти кости самостоятельны.
Проксимальные концы обеих костей образуют сочленовную ямку для соединения с плечом; позади этой ямки находится локтевой отросток (рис. 61, 6) локтевой кости, ограничивающий разгибание конечности.
Запястье (carpus, рис. 61, 7) состоит из двух рядов мелких костачек. К дистальному ряду косточек запястья примыкают пять удлиненных косточек пясти (metacarpus; рис. 61, 8). С дистальными концами пястных костей сочленяются фаланги пальцев (phalanges digitorum; рис. 61, 9). У земноводных первый (большой) палец сильно редуцирован и кисть заканчивается лишь четырьмя хорошо развитыми пальцами.
Рис. 61. Передняя конечность и плечевой пояс лягушки: 1 — плечевая кость, 2 — головка плеча, 3 — предплечье, 4 — локтевая кость, 5 — лучевая кость, 6 — локтевой отросток, 7 — запястье, 8 — пясть, 9 — фаланги пальцев, 10 — лопатка, 11 — надлопаточный хрящ, 12 — коракоид, 13 — суставная впадина для головки плеча, 14 — прокоракоидный хрящ, 15 — ключица, 16 — грудина, 17 — предгрудинник, I — редуцированный первый палец, II — V — хорошо развитые пальцы
Пояс передних конечностей, или плечевой пояс, у земноводных, как и у акуловых рыб, лежит в толще мускулатуры туловища, связывающей его с осевым скелетом. Из верхней (дорзальной) лопаточной части первичного пояса образуется лопатка (scapula; рис. 61, 10); его самая верхняя часть остается хрящевой в виде широкого надлопаточного хряща (cartilago suprascapularis, рис. 61, 11). На передне-наружной поверхности надлопаточного хряща у некоторых бесхвостых амфибий имеется небольшое окостенение — остаток клейтрума рыбообразных предков. Окостеневшая коракоидная часть пояса превратилась в мощную коракоидную кость (coracoideum; рис. 61,12), вместе с лопаткой образующую суставную впадину для головки плеча (рис. 61, 13). Кпереди от коракоида за небольшим отверстием лежит хрящевой прокоракоид (cartilago procoracoidea; рис. 61, 14), на который налегает тоненькая покровная кость — ключица (clavicula; рис. 61, 15). Неокостеневшие хрящевые внутренние концы коракоидов и прокоракоидов правой и левой сторон сливаются вместе по средней линии. Позади коракоидов располагается костная грудина (sternum, рис. 62, 16) с хрящевым задним концом. Впереди от прокоракоидов выдается предгрудинник (praesternum; рис. 61, 17) также с хрящевым концом. В поясе передних конечностей хвостатых земноводных заметно больше хряща, а окостенения имеют меньшие размеры; ключицы часто не развиваются.
Плечевой пояс служит опорой для передних конечностей и местом прикрепления управляющих ими мышц.
Грудная клетка у земноводных не развивается: грудина не сочленяется с ребрами.
Рис. 62. Задняя конечность (А) и тазовый пояс (Б) лягушки сбоку: 1 — бедренная кость, 2 — головка бедра, 3 — голень, 4 — большая берцовая кость, 5 — малая берцовая кость, 6 — предплюсна, 7 — tibiale, 8 — fibulare, 9 — плюсна, 10 — фаланги пальцев, 11 — рудимент VI пальца, 12 — подвздошная кость, 13 — седалищная кость, 14 — лобковый хрящ, 15 — вертлужная впадина, I — V — пальцы
Проксимальный ряд костей предплюсны (tarsus, рис. 62, 6) бесхвостых земноводных состоит из двух удлиненных костей, образующих добавочный рычаг конечности. Внутренняя из них называется tibiale (astragalus; рис. 62, 7; примыкает к большеберцовому краю голени), наружная — fibulare (calcaneus, рис. 62, 8). Между голенью и этими костями образуется голеностопный сустав. От дистального ряда костей предплюсны у земноводных сохраняются лишь 2 — 3 маленькие косточки. Плюсна (metatarsus; рис. 62, 9) образована пятью длинными косточками, к которым причленяются фаланги пальцев (phalanges digitorum; рис. 62, 10). Самый длинный палец у лягушек — IV. Сбоку от I (внутреннего) пальца располагается маленький рудимент VI («предпервого») пальца (praehallus; рис. 62, 11).
Пояс задних конечностей, или тазовый, у земноводных, как и у всех наземных позвоночных, состоит из трех парных элементов; причем все они вместе образуют суставную вертлужную впадину (acetabulum; рис. 62, 15) для соединения с головкой бедра. Длинные, направленные вперед подвздошные кости (ilium; рис. 62, 12) своими концами причленяются к поперечным отросткам крестцового позвонка (см. рис. 54). Нижняя часть тазового пояса у земноводных не окостеневает и представлена лобковым хрящом (cartilago pubis, рис. 62, 14). Позади него лежат парные седалищные кости (ischium; рис. 62, 13).
У хвостатых земноводных, по сравнению с бесхвостыми, в тазовом поясе много больше хряща, а сформировавшиеся кости малы.
ЗАКЛЮЧЕНИЕ
Земноводные (Amphibia) — первый класс наземных позвоночных животных. Однако у представителей класса еще сохраняется постоянная связь с водой. Эта двойственность отчетливо проявляется в особенностях эмбрионального и постэмбрионального развития. Яйцеклетки (икра) могут развиваться только в воде (или в редких случаях в предельно влажной среде). Из яйца выходит личинка — головастик, имеющая отчетливо выраженные признаки типичного водного животного: у него есть жабры и поддерживающие их жаберные дуги, сердце двухкамерное, один круг кровообращения, парные конечности наземного типа отсутствуют, основной орган движения — мощный хвостовой плавник, развиты органы боковой линии и т. п. Во время метаморфоза (превращения) личинка теряет часть признаков, свойственных водным животным, и приобретает особенности, обеспечивающие переход к наземному (вернее, наземно-водному) образу жизни: появляются парные конечности наземного типа, развиваются легкие, редуцируются жабры и поддерживающий их скелетный аппарат, перестраивается кровеносная система — образуются два неполностью разделенных круга кровообращения и т. п. Двойственность организации как приспособление к наземно-водному образу жизни хорошо выражена и у взрослых особей. Наземный образ жизни обеспечивается рядом особенностей строения : большая дифференцировка позвоночного столба на отделы и более прочное соединение тел позвонков друг с другом (замена амфицельных позвонков процельными или опистоцельными); появление парных конечностей наземного типа; усложнение строения и большая прочность поясов конечностей (при этом уже устанавливается достаточно прочное соединение тазового пояса с осевым скелетом); сильная редукция ме-тамерной мускулатуры и ее замена достаточно мощным сложным комплексом мускулов; появление век (защита глаз от механических повреждений, предотвращение подсыхания роговицы и т. п.); образование полости среднего уха с барабанной перепонкой и слуховой косточкой — стремечком (обеспечение возможности слуха в воздушной среде). Большую роль сыграло исчезновение жабр и развитие легких, гортани и хоан, создающих возможность воздушного дыхания; возникновение двух кругов кровообращения; большая дифференцировка пищеварительной системы (большие энергетические затраты при поддержании тела в воздушной среде) и т. п. Общее удлинение задних конечностей, обособление в них дополнительного рычага (благодаря резкому удлинению двух проксимальных косточек предплюсны) и возможность сильного перегиба посредине туловища в месте сочленения ветвей подвздошных костей с поперечными отростками крестцовых позвонков — приспособления к движению прыжками у бесхвостых земноводных. У ползающих хвостатых земноводных этих особенностей нет. Слияние в единое целое двух костей предплечья и двух костей голени связано с резким уменьшением необходимости вращательных движений стопы и кисти при перемещении прыжками. У хвостатых земноводных как предплечье, так и голень состоят из двух самостоятельных элементов, обеспечивая необходимые при ползании вращательные движения кисти и стопы. «Водные» черты строения проявляются в целом ряде особенностей: относительно слабое развитие окостенений скелета, обилие слизистых желез в коже (покрывающая кожу слизь уменьшает трение при движении в воде, предотвращает проникновение в кожу бактерий и грибков и т. п.), сохранение хвоста, часто уплощенного с боков и окаймленного кожистой складкой (тритоны и другие хвостатые земноводные), большое сходство мочеполовой системы с большинством групп рыб, характерное для подавляющего большинства видов земноводных наружное оплодотворение и т. д. При относительно небольшой поверхности легких земноводных необходимы достаточно мощные дополнительные органы дыхания. Таким органом становится всегда влажная (благодаря обилию слизистых желез), легко проницаемая для влаги и газов кожа и отчасти слизистая оболочка ротовой полости. У прудовой лягушки в активном состоянии легкие поглощают до 50% потребляемого организмом кислорода и выделяют только около 14% углекислого газа; путем кожного дыхания поглощается до 50% кислорода и выделяется до 86% углекислого газа. У ведущей более наземную жизнь травяной лягушки при легочном дыхании поступает до 67% кислорода и выделяется до 26% углекислого газа, а путем кожного дыхания поглощается 33% кислорода и выделяется 74% углекислого газа. При возрастании уровня метаболизма (усиление общей активности и всех процессов обмена при повышении температуры среды) удельная роль легких в обеспечении организма кислородом заметно возрастает. Понижение температуры среды вызывает снижение уровня метаболизма. При этом кожное дыхание почти в полной мере обеспечивает и насыщение организма кислородом и выделение углекислого газа, а относительное значение легких в дыхании резко снижается. Эта двойственность характера дыхания объясняется не только недостаточным развитием поверхности легких и несовершенством легочной вентиляции («заглатывание» воздуха при отсутствии грудной клетки); она необходима при земноводном образе жизни представителей этого класса. Именно эта двойственность органов дыхания обеспечивает земноводным возможность длительного пребывания в воде (вплоть до зимовки на дне водоема многих видов бесхвостых, когда при резком снижении уровня метаболизма кожное дыхание полностью обеспечивает все потребности организма в кислороде и выделении углекислого газа). Использование кожи для дыхания возможно лишь тогда, когда она легко проницаема для влаги и газов. Но такая кожа не может предохранять организм от больших потерь воды (высыхания). Поэтому практически почти все виды земноводных населяют лишь сырые, влажные участки, где организм теряет меньше влаги и всегда может восполнить ее потерю. У относительно мало связанных с водой жаб (зимуют на суше, в водоемы уходят только для икрометания) кожа уплотнена; это снижает возможности кожного дыхания, что компенсируется увеличением внутренней поверхности легких. Однако и у них, несмотря на уплотнение кожи, за ночной период охоты организм теряет до 15—30% воды. Некоторому уменьшению влагопотерь (при сохранении проницаемости кожи) у земноводных помогает слизь, покрывающая кожу.
Обширные подкожные лимфатические полости служат резервуарами запасной воды. Уменьшаются влагопотери и благодаря обратному всасыванию воды в мочевом пузыре, заднем отделе кишечника и в клоаке. Очень резко уменьшаются влагопотери благодаря приспособительным особенностям поведения: повышенную активность земноводные проявляют только в часы максимальной влажности воздуха (в ясную погоду — в сумерках, а также ночью), на отдых они уходят в норки, где за счет почвенной влаги поддерживается высокая влажность. Двойственность органов дыхания делает невозможным полное разделение большого и малого кругов кровообращения. Но специфические особенности в строении сердца и отходящих от него кровеносных стволов обеспечивают некоторое разобщение кровотока несмотря на то, что в сердце земноводных только один желудочек, а в верхних полых венах есть примесь артериальной крови. Развитие мускульных выростов стенок желудочка уменьшает перемешивание крови, а от-хождение артериального конуса от правой (более венозной) стороны желудочка и детали его внутреннего строения (последовательность отхождения артериальных дуг, устройство спирального клапана) позволяют направлять более венозную кровь в кожу и легкие, более артериальную — к головному мозгу и органам чувств. Большая, по сравнению с рыбами, дифференцировка пищеварительного тракта приводит к некоторому повышению интенсивности использования пищи. Однако скорость пищеварения у земноводных невелика и зависит от температуры окружающей среды. Пищевые связи довольно простые; круг используемых кормов невелик (только животные относительно мелких размеров). Земноводные, как и рыбы, характеризуются непостоянством температуры тела (пойкилотермность): у земноводных она обычно лишь на 0,5—1°С выше температуры окружающей среды. Лишь в период наивысшей активности (преследование добычи, уход от опасности) температура тела может превышать температуру среды на 5—7°С.
Пойкилотермность вызывает у земноводных умеренных и северных широт резко выраженную сезонную смену активности: при снижении температуры воздуха до +5 — +8°С все земноводные уходят в укрытия (часть видов лягушек — в ямы на дне водоемов; большинство видов бесхвостых и хвостатых земноводных скрываются в норах грызунов, прогнивших корнях деревьев, кучах мха и т.п.) и впадают в состояние оцепенения. С этим связано и географическое распространение земноводных: большинство видов этих животных свойственны тропической зоне. В тропиках, при относительно устойчивых в течение года температурных условиях, у ряда видов земноводных состояние оцепенения вызывается резким снижением влажности воздуха («спячка» в сухой период года). Очень большая зависимость земноводных от влажности и температуры среды сказывается в том, что погодные условия (в наших широтах — сильные засухи летом, сильные морозы без снега зимой) часто служат основной причиной смертности и определяют резкие колебания численности земноводных по годам.
Дополнительная литература
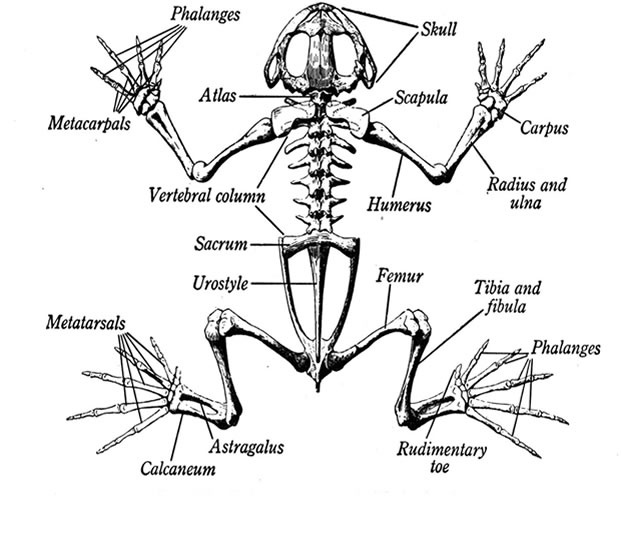
Скелет земноводных, как и у всех позвоночных, состоит из черепа, позвоночника, скелета конечностей и их поясов. Череп почти сплошь хрящевой. Он подвижно сочленен с позвоночником.
Позвоночник содержит девять позвонков, объединенных в три отдела: шейный (1 позвонок), туловищный (7 позвонков), крестцовый (1 позвонок), а все хвостовые позвонки срослись, образовав единую косточку — уростиль. Ребра отсутствуют. Плечевой пояс включает типичные для наземных позвоночных кости: парные лопатки, вороньи кости (коракоиды), ключицы и непарную грудину. Он имеет вид полукольца, лежащего в толще туловищной мускулатуры, т. е. не соединен с позвоночником.
Тазовый пояс образован двумя тазовыми костями, образованными тремя парами подвздошных, седалищных и лобковых костей, сросшихся между собой.
Длинные подвздошные кости причленены к поперечным отросткам крестцового позвонка.
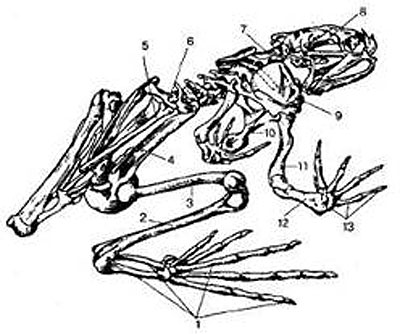 |
1 — кости стопы; 2 — голень; 3 — бедренная кость; 4 — подвздошная кость; 5 — уростиль; 6 — крестцовый позвонок; 7 — шейный позвонок; 8 — череп; 9 — лопатка; 10 — грудина; 11 — плечевая кость; 12 — предплечье; 13 — кости кисти. |
Скелет свободных конечностей построен по типу системы многочленных рычагов, подвижно соединенных шаровидными суставами. В составе передней конечности выделяют плечо, предплечье и кисть. У бесхвостых земноводных локтевая и лучевая кости сливаются, образуя общую кость предплечья. Кисть подразделяется на запястье, пясть и четыре фаланги пальцев. Задняя конечность состоит из бедра, голени и стопы. Стопа включает кости предплюсны, плюсны и фаланги пяти пальцев. Задние конечности длиннее передних. Это связано с передвижением по суше прыжками, а в воде — с энергичной работой задних конечностей при плавании. Как видим, такая схема строения конечностей является типичной для наземных позвоночных и в каждом классе имеет незначительные изменения, связанные с особенностями их движения. Благодаря подвижности отделов скелета движения тела земноводных более разнообразны, чем у рыб.
К этому можно добавить, что именно эксперименты на лягушках позволили установить основные закономерности функционирования нервов, мышц, сердца, гладких мышц, и многих других органов и тканей.
Сейчас лягушка по-прежнему является основным объектом, на которых исследователи проводят свои первые опыты.
Самыми распространенными заданиями для начинающих биологов являются, пожалуй, исследования работы сердца лягушки. Протоколы и процедуры опытов описаны в тысячах руководств и практикумов на всех языках мира. Однако из-за отсутствия надлежащих навыков экспериментатор вначале видит не совсем то, что нарисовано в пособиях.
Мы предлагаем для просмотра и изучения несколько реальных фотографий и видеоклипов, поясняющих особенности строения и функционирования сердца лягушки. Все записи проведены в течение одного эксперимента на лягушке Rana temporaria.
Внешнее строение сердца.
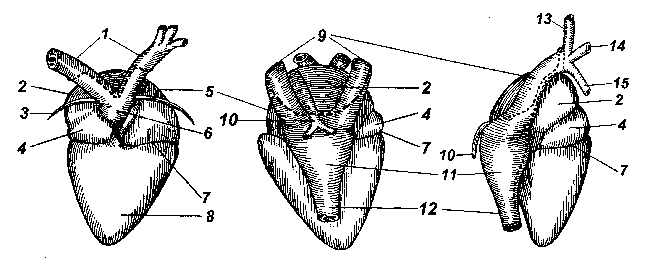
Рис. 1
Сердце у лягушек расположено в грудобрюшной полости под грудиной. Его общее строение с дорсальной и вентральной сторон и сбоку (справа) показано на Рис. 1: 1 — дуги аорты; 2 — правое предсердие; 3 — щетинка, проведенная в отверстие под артериальным стволом; 4 — луковица аорты (артериальный конус); 5 — левое предсердие; 6 — артериальный ствол; 7 — венечная борозда; 8 — желудочек; 9 — передние (краниальные) полые вены; 10 — легочные вены (на виде сбоку только правая вена); 11 — венозный синус; 12 — задняя (каудальная) полая вена; 13 — наружная яремная вена; 14 — безымянная вена; 15 — подключичная вена).
Традиционно считается, что сердце земноводных состоит из трех камер, двух предсердий и одного желудочка. Строго говоря, это не совсем так. К сердцу относятся еще два отдела, явственно обособленные как отдельные камеры у низших позвоночных: рыб, амфибий, некоторых рептилий. Это венозный синус (венозная пазуха) и артериальный конус (луковица аорты).
Венозный синус представляет собой тонкостенную камеру, образованную слиянием полых вен — задней (каудальной) и двух передних (краниальных), левой и правой. Синус располагается с дорсальной стороны сердца, и его можно рассмотреть, если аккуратно оттянуть верхушку сердца вперед, к голове.
Мускулистый артериальный конус расположен вентрально между желудочком и коротким артериальным стволом (он является частью сосудистой системы), от которого отходят левая и правая дуги аорты. Артериальный ствол не прирастает к вентральной поверхности предсердий, под него можно провести тонкий волосок (щетинка «3» на Рис.1).
Мышечные стенки венозного синуса и артериального конуса сокращаются, и в какой-то степени участвуют в перемещении крови.
Однако внешне артериальный конус не отличается от крупного сосуда. Между полыми венами и венозным синусом также нет четкой границы, нельзя определенно указать, где заканчиваются полые вены и начинается венозный синус.
Почему венозный синус и артериальный конус считают частью сердца, а не частью сосудистой системы? Это сделано на основании особенностей эмбрионального развития сердца и сосудов. В ходе эмбриогенеза сердце, включающее венозный синус, предсердие(-я), желудочек(-чки), артериальный кону, возникает из одного зачатка, а сосуды — из другого. У теплокровных животных на определенной стадии развития венозный синус и артериальный конус также отчетливо выражены. Затем венозный синус трансформируется в синусный узел (пейсмекерную зону, пейсмекер), расположенный в стенке правого предсердия, а артериальный конус преобразуется в безмышечное кольцо, расположенное на границе между левым желудочком и аортой. Таким образом, синусный узел теплокровных животных является гомологом венозного синуса низших позвоночных.
Пейсмекер (англ. «pacemaker», водитель ритма) определяет ритм сердечных сокращений. Здесь имеются специализированные мышечные волокна, обладающие автоматией. В пейсмекерных кардиомиоцитах спонтанно, с определенной периодичностью, возникают волны возбуждения, которые затем последовательно распространяются на миокард предсердий, желудочка и артериального конуса. На границах различных отделов сердца (у лягушки между венозным синусом и предсердиями, предсердиями и желудочком, желудочком и артериальным конусом) волна возбуждения проводится с меньшей скоростью, возникает задержка проведения возбуждения, сопровождаемая такой же задержкой волны сокращения.
Венозный синус сердца лягушки сообщается с правым предсердием через широкое овальное отверстие, окруженное мускулистым синоатриальным кольцом. Сокращения синоатриального кольца частично препятствуют обратному току крови из правого предсердия в венозный синус. Других клапанных структур здесь нет.
Легочные вены, несущие аэрированную кровь, перед входом в левое предсердие объединяются в общую легочную вену. Настоящих клапанов здесь также нет.
Передние полые вены образуются слиянием наружных яремных, подключичных и безымянных вен. Короткая задняя полая вена выходит из печени.
Предсердия отделяются от желудочка венечной бороздой. Она разделяет сердце на переднюю часть (предсердия, входящие и выходящие сосуды), и заднюю часть (желудочек).
Снаружи сердце окружено перикардом, который можно представить себе как тонкостенный мешочек, надвинутый на сердце со стороны его верхушки. Внутренний листок перикарда (или эпикард) является наружным слоем сердца. Между эпикардом и наружным листком перикарда, в перикардиальной полости, находится перикардиальная жидкость. Граница прикрепления наружного листка перикарда к стенкам сердца и сосудов показана на Рис.1 пунктирной линией.
В сердце лягушки коронарные сосуды имеются только в стенках артериального конуса. Ткани венозного синуса, предсердий и желудочка снабжаются кислородом за счет перекачиваемой крови.
Миокард предсердий не переходит непосредственно в миокард желудочка. Контакт между ними осуществляется через относительно компактный пучок специализированной мышечной ткани, расположенный в области атриовентрикулярного отверстия, являющегося общим входом для правого и левого предсердия. Здесь имеются хорошо выраженные атриовентрикулярные створчатые клапаны.
Исследование работы сердца лягушки in situ
|
Рис. 2 |
У всех животных сердце хорошо работает при оптимальном уровне внутрисердечного давления. Большая кровопотеря, приводящая к снижению кровенаполнения сердца, приводит к уменьшению силы сердечных сокращений. Поэтому препаровку следует проводить аккуратно, не повреждая крупные сосуды. |
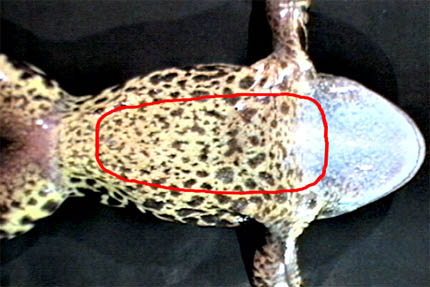
После удаления кожного лоскута ножницы и пинцет следует промыть водой и вытереть.
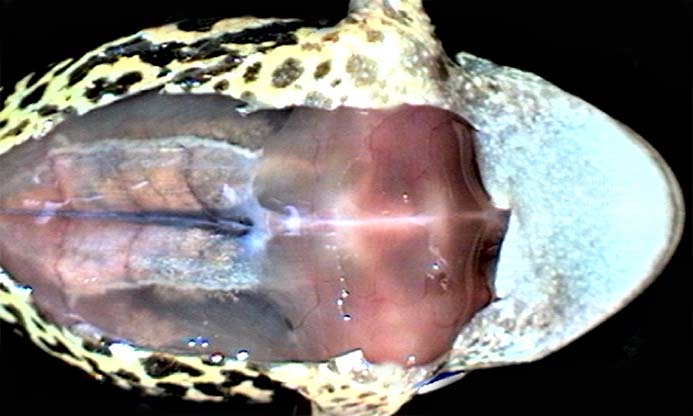
Рис. 3
Среди различных мышц на брюшной поверхности хорошо выделяется прямая мышца живота (Рис. 3). По оси тела отчетливо видна брюшная вена. Если навести курсор на область зоны с удаленной кожей, появится схема (проекция) костей плечевого пояса. Небольшим пинцетом сбоку от брюшной вены захватывают и приподнимают кверху край хрящевой части грудины (os sternum). Вскрывают мускульную стенку грудобрюшной полости рядом с пинцетом кончиками острых ножниц. ОСТОРОЖНО: брюшная вена рядом с каудальным краем грудины уходит вертикально вниз (для наблюдателя), и нужно постараться не перерезать ее. Не опуская край грудины, разрез продолжают вдоль нее слева и справа до врановых костей (os coracoideum). Затем большими ножницами перерезают врановые кости, ключицы (claviculae), продолжают разрез до передней части предгрудины (episternum), и удаляют весь этот мышечно-костный лоскут. После этого передние лапки и край нижней челюсти лягушки перекалывают так, чтобы расширить операционное поле.
|
С помощью тонкого пинцета и острых ножниц удаляют париетальный листок перикарда с вентральной поверхности сердца, после чего становятся хорошо видимыми желудочек, вентральные участки предсердий, луковица и дуги аорты (Рис. 4). У летних и осенних лягушек венечная борозда (граница между предсердиями и желудочком) часто заполнена желтовато-коричневой жировой тканью, в которой проходят кровеносные сосуды. В стенках предсердий под бинокулярным микроскопом хорошо различимы светлые тяжи — пучки мышечных волокон на внутренней поверхности предсердий (трабекулы). С трех сторон сердце окружено долями печени, в каудальной части операционного поля хорошо виден синеватый желчный пузырь. Рис. 4 |
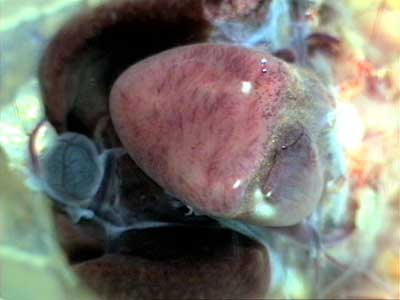 |
 |
При бoльшем увеличении в эпикарде видны многочисленные черные отростчатые клетки — меланоциты (Рис. 5). Такие клетки можно наблюдать по ходу многих сосудов. Например, общее устье легочных вен при небольшом увеличении микроскопа выглядит практически черным из-за скоплений меланоцитов. Меланоциты определяют общую окраску кожи лягушки. В лабораторных условиях изменения окраски кожи лягушки являются хорошими индикаторами функционального состояния животных. Низкая влажность, чрезмерное освещение, голодание, высокая температура (для наших лягушек более +20оC) приводят к посветлению лягушек. При высокой влажности и низкой температуре кожа лягушки темнеет. Функциональное значение меланоцитов в эпикарде для работы сердца не изучено. |
Рис. 5
Включение видеоклипа позволит вам наблюдать последовательные сокращения предсердий и желудочка (Клип 1). Обратите внимание на отчетливую временную задержку между систолой предсердий и желудочка. Такая задержка характерна для сердец всех животных, имеющих камерные сердца (позвоночные и моллюски). Атриовентрикулярная задержка повышает эффективность работы сердца, т.к. в этот период большая часть крови из предсердий успевает переместиться в желудочек.
In situ аортальные стволы оттягивают основание сердца дорсально. Поэтому для рассмотрения околосердечных сосудов вблизи дорсальных участков предсердий следует слегка оттянуть желудочек в каудальном направлении с помощью серфинки. Серфинка представляет собой легкий зажим, выполненный из упругой проволоки. С ее помощью аккуратно захватывают верхушку желудочка. Продольная ось серфинки должна совпадать с продольной осью сердца. К кольцу серфинки предварительно привязывают лигатуру (нить), с помощью которой экспериментатор соединяет сердце с кимографом и другими датчиками.
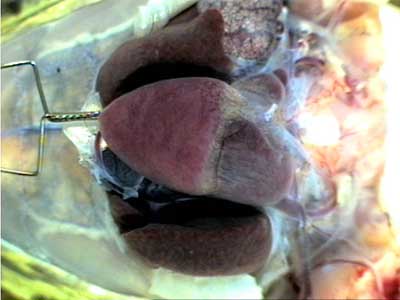
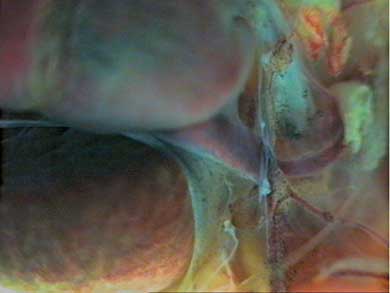
Небольшое растяжение сердца в кранио-каудальном направлении (влево на Рис. 6) позволяет лучше рассмотреть элементы вентральной аорто-артериальной системы, и увидеть некоторые участки венозной части сердца (Рис. 6). Становятся отчетливо видимыми оба предсердия, луковица аорты, аортальный ствол, дуги аорты, а ниже, в глубине, видны основания передних полых вен. Некоторые особенности строения венозного русла можно рассмотреть на следующем рисунке при бoльшем увеличении (Рис. 7). В левом верхнем квадранте рисунка проецируется часть левого предсердия, оно не в фокусе видеокамеры, а внизу видны основные вены, формирующие левую переднюю (краниальную) полую вену.
|
Рис. 6 |
Рис. 7 |
На видеоклипах, сопровождающих эти фотографии (Клип 2, Клип 3) следует обратить внимание на последовательность сокращений полых вен, предсердий, желудочка и артериального конуса.
Эти наблюдения показывают, что в сердце лягушки возбуждение и сокращение первоначально возникают в области полых вен, затем происходит систола предсердий, желудочка и артериального конуса.
Наблюдение нормальных сокращений сердца показывает, что в данный момент источником сердечного ритма (пейсмекером) является участок венозного синуса, примыкающий к левой полой вене (Клип 3).
|
Для исследования распространения активности в первичной пейсмекерной зоне, включающей устья полых вен и венозный синус, следует провести еще одно оперативное вмешательство. Если мы оттянем верхушку желудочка вперед, к голове, мы увидим дорсальную поверхность предсердий и желудочка (Рис. 8). Оказывается, что примерно в верхней трети дорсальная часть желудочка соединена с перикардом основания сердца тонким соединительнотканным жгутом — уздечкой сердца. Остатки перикарда и уздечка выглядят здесь как голубоватые полупрозрачные соединительнотканные пленочки. Рис. 8 |
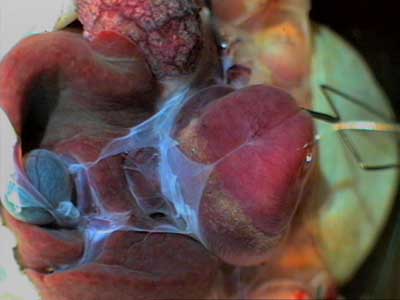 |
На видеоклипе (Клип 4) видно, что уздечка существенно ограничивает движения желудочка in situ. При любых способах регистрации сокращений сердца лягушек in situ следует перерезать уздечку сердца. Это является необходимым условием для успешного проведения эспериментов.
После удаления уздечки и остатков перикарда между предсердиями и полыми венами область венозного синуса становится отчетливо различимой (Рис. 9). Справа она хорошо выявляется после смещения желудочка влево по отношению к оси тела животного (на рисунке — вниз) (Рис. 10). Вены заполнены кровью, и их границы легко уставливаются. Задняя полая вена очень короткая, об этом нужно помнить при проведении экспериментов. Лигатуры и перерезки при выделении сердца следует производить ближе к печеночному краю вены, иначе можно повредить венозный синус, и изолированное сердце перестанет сокращаться.
|
Рис. 9 |
Рис. 10 |
Обратите внимание на светлое кольцо на границе венозного синуса и правого предсердия. Это мышечное кольцо (синоатриальное кольцо, Рис. 9) расположено между двумя камерами (венозным синусом и правым предсердием), активно сокращается, и частично выполняет функцию клапана.
Фазы сокращений различных участков венозного синуса с правой стороны сердца можно проанализировать на (Клип 5). В нормальных условиях последовательность распространения возбуждения зависит от состояния полых вен, венозного синуса и близкорасположенных участков сердца.
Зоны первичной электрической активности, определяющих ритм сердечных сокращений, могут перемещаться при изменении функционального состояния сердца. Это происходит и в норме, и при ухудшении внешних условий. Длительное переживание в условиях in vitro, согревание сердца лягушки более чем до 20 oC приводит к появлению аритмий. Сначала нарушается проведение возбуждения из предсердий в желудочек, а затем происходят только сокращения полых вен и венозного синуса. Последовательность сокращений в этой зоне в норме вы можете проанализировать на Клипе 6. Следующий клип (Клип 7) показывает изменения работы венозного синуса и предсердий через 15 минуты. Произошел полный синоатриальный блок, волна возбуждения не проходит в предсердия, место блока — синоатриальное кольцо.
Литература
Дыхательная система земноводных представлена легкими и кожей, через которую они также способны дышать. Легкие – это парные полые мешки, имеющие ячеистую внутреннюю поверхность, которая усеяна капиллярами. Именно здесь и происходит газообмен. Механизм дыхания лягушек относится к нагнетательным и его нельзя назвать совершенным. Лягушка набирает воздух в ротоглоточную полость, что достигается путем опускания дна ротовой полости и открывания ноздрей. Потом дно ротовой полости поднимается, и ноздри снова закрываются клапанами, а воздух нагнетается в легкие.
Кровеносная система лягушки состоит из трехкамерного сердца (два предсердия и желудочек) и двух кругов кровообращения – малого (легочного) и большого (туловищного). Малый круг кровообращения у земноводных начинается в желудочке, проходит через сосуды легких и заканчивается в левом предсердии.
Большой круг кровообращения начинается тоже в желудочке, проходит по всем сосудам тела земноводного, возвращается в правое предсердие. Как и у млекопитающих, кровь насыщается в легких кислородом, а затем разносит его по всему телу. В левое предсердие попадает артериальная кровь из легких, а в правое – венозная кровь из остального тела. Также в правое предсердие попадает кровь, которая проходит под поверхностью кожи и насыщается там кислородом.
Несмотря на то, что в желудочек попадает и венозная, и артериальная кровь, она там не смешивается полностью благодаря наличию системы клапанов и карманов. Из-за этого артериальная кровь идет к мозгу, венозная – направляется к коже и в легкие, а смешанная – к остальным органам. Именно из-за наличия смешанной крови интенсивность жизненных процессов земноводных невысокая, а температура тела часто может меняться.